![]()
Э.А.Мирзоян
ТЕОРИЯ, ПРАКТИКА И ОПЫТ
Философский анализ
Мозжечковые связи
Афферентные связи
Анализируя главы последнего труда академика В.В.Фанарджяна, посвящённые мозжечку, приходишь к очень интересным выводам относительно целого и частного в головном мозге. Как известно, существует соматотопическое распределение в отдельных долях мозжечка по приоритету в управлении различных зон человеческого организма. То же самое можно сказать и о других областях головного мозга.
Так, например, в ростральной области мозжечка расположены следующие зоны представительства тела человека.
В передней зоне – область ног, чуть ниже, к середине – само туловище, а внизу – шея и голова. В каудальной части мозжечка вся картина построена с диаметральной противоположностью.
Вверху каудального отдела расположено представительство головы и шеи, а внизу – ног, туловище, вновь – посередине. При этом необходимо учитывать также и билатеральность органа.
Из вышесказанного вытекает два вывода:
1. Как афферентная, так и эфферентная системы мозжечка «строят» голографическую картину; т.е. импульсация не просто подаётся в мозжечок в виде потенциалов и таким же образом, как считается ныне, выходит из мозжечка, но поступая в цифровом виде, в той или иной музыкальной транспозиции, ритме и тоне, отображается уже в самом мозжечке как объёмными и, что немаловажно, целостными голограммами.
2. Если же исходить из закона «Всё или ничего», то импульсы передаются с наиболее полной силой. Отсюда следует, что идеации, которые несут сами импульсы, также максимально полные. Таким образом, импульсации передают идеации в мозжечке, да и во всём головном мозге, где они выявляются в тех или иных областях в виде голограмм.
Мы ещё вернёмся к вопросу о преобразовании идеаций в мозге в голограммы, теперь отступим, перейдя к крупным мозжечковым связям.
По выражению князя Орбели, мозжечок является отделом, который регулирует и стабилизирует все рефлекторные дуги в целом. И, как показала в своих экспериментах лаборатория Л.А.Орбели, при удалении мозжечка у животных происходило бесконтрольное высвобождение рефлексов, те же аффекты наблюдались у человека при патологиях мозжечка. Ссылаясь на труды академика Л.А.Орбели, В.В.Фанарджян писал:
«Анализируя все эти расстройства – атонию, атаксию, астазию, асинергию и др., академик Л.А.Орбели (1935,1940) пришёл к выводу, что несчастье безмозжечковых животных заключается в том, что не то, что отсутствуют, а слишком неуклонно и хорошо осуществляются лабиринтные, проприоцептивные и другие рефлексы». Если провести небольшой философский анализ вышеизложенного, то вывод очевиден: мозжечок – это Воля головного мозга, постоянно направленная на фактор сдерживания и сбалансирования всех импульсаций и идеаций. Так функционирует печень в организме.
Кроме лазящих и мшистых волокон мозжечок связан с отделами мозга посредством собственных ядер.
Ядра мозжечка
Философское рассмотрение взаимосвязи мозжечка и таламуса
Рассмотрев нейроны коры мозжечка и проведя анализ тех функций, которые присущи им, следует сейчас подойти к рассмотрению ядер мозжечка и их организации. Перечисление ядер мозжечка:
1. Фастигиальное ядро мозжечка;
2. Промежуточное (интерпозитарное) переднее ядро, пробковидное;
3. Промежуточное (интерпозитарное) заднее ядро, шаровидное;
4. Латеральное (зубчатое) ядро.
Ядра мозжечка являются одними из важнейших центров интеграции и организации процессов в ЦНС. Они являются первой ступенью в интегративной программе всего мозжечка. В данном подразделе будут объяснены, с точки зрения философского аспекта, опосредованная и непосредственная роли ядер мозжечка в интегративной и координирующей функциях связанных с ЦНС.
Ядра мозжечка получают афферентацию почти от всех отделов ЦНС, окончательно проводят интеграцию информационных субстратов и отправляют в те же отделы (реципрокность), а также и к коре, как отделу собирательному и отражательному.
Центральные ядра располагаются в медиолатеральном направлении, в белом веществе. Наибольшее развитие, как отмечает Аматуни, центральные ядра (ЦЯ) получили у млекопитающих, но впервые появились у амфибий и рептилий; из них самое древнее является фастигиальное ядро. Как и все ядерные образования в мозге, фастигиальное ядро имеет сложную архитектонику и состоит из крупноклеточной и мелкоклеточной частей. Нейроны обеих частей – радиального строения и отличаются между собой, в морфологическом аспекте, ветвистостью и величиной дендритов и аксонов. Кроме того, как указывают исследования конца прошлого столетья, наряду с афферентными волокнами из различных структур мозга, а также и от клеток Пуркинье коры мозжечка, нейроны фастигиального ядра (аксоны средних клеток) имеют соединения с противоположным ФЯ. Те же связи наличествуют и в других билатеральных структурах.
Прежде, чем начать говорить о роли как фастигиального ядра, так и остальных центральных ядер мозжечка, необходимо сказать, что образования в ЦНС, имеющие множества афферентных входов от различных образований и эфферентных выходов, не могут быть носителями одной локальной функции, но играют роль носителей интеграционной функции, с той или иной доминирующей иерархической окраской. Для того, чтобы представить хотя бы общую картину взаимоотношений между ЦЯ мозжечка и другими отделами ЦНС, необходимо представить перечень известных и предполагаемых связей самих структур с ядрами мозжечка, не нарушая при этом самого основного положения Природы - Иерархии. Итак, перейдём к иерархическим взаимосвязям в фастигиальном ядре и начнём с таламуса. В связи с этим интересны морфологические исследования Мелик-Мусян, в частности, её статья за 1979г. «К особенностям мозжечково-таламических связей». В данной статье автор указывает на следующие связи между мозжечковыми ядрами и ядрами таламуса, выявленные методом дегенерации. Ниже представлена схематическое представительство ядер таламуса в центральных ядрах мозжечка. Несмотря на то, что каждое ядро мозжечка имеет множество связей от различных отделов ЦНС, на схеме представлены несколько связей с таламическими ядрами:
1. Фастигиальное ядро – вентрально-латеральное ядро таламуса;
2. Фастигиальное ядро – вентрально-медиальное ядро таламуса;
3. Фастигиальное ядро – вентро-постерио-медиальное ядро таламуса;
4. Промежуточные ядра – вентрально-латеральное ядро таламуса;
5. Промежуточные ядра – вентро-постерио-медиальное ядро таламуса;
6. Промежуточные ядра – вентро-постерио-латеральное ядро таламуса;
7. Латеральное (Зубчатое) ядро – вентрально-латеральное ядро таламуса;
8. Латеральное (Зубчатое) ядро – вентрально-антериорное ядро таламуса;
9. Латеральное (Зубчатое) ядро – вентро-постерио-латеральное ядро таламуса;
10. Латеральное (Зубчатое) ядро – вентро-постерио-медиальное ядро таламуса;
11. Латеральное (Зубчатое) ядро – вентрально-медиальное ядро таламуса.
Теперь, не останавливаясь на самих связях таламических центров с мозжечковыми, вновь подойдём к рассмотрению скрытых аспектов, как таламуса, так и мозжечка.
Все ядра таламуса, до их активации, представляют собой 4-й принцип – Кама Манас. Так как все 42 ядра, по своей сути, являются релейными центрами, то и сам таламус отвечает за компараторно-релейную принципацию в мозге. По всей видимости, таламус также действует в эзотерическом аспекте; высшие вибрации сопоставляются с низшими, а в результате выдаётся оптимальное значение. Таким образом, централизованный аспект таламуса проявляется на всех планах.
Старое название – зрительные бугры, подтверждающееся тем, что таламус один из первичных зрительных центров - также можно принять и в виде метафоры, т.к. зрение тоже несёт роль сопоставителя.
Таламус входит в межуточный мозг и его по праву можно назвать внутренним мозгом в мозге. По своей иерархической градации таламус соответствует Манасу, причём до активности эпифиза таламус подчинён вибрациям Низшего Манаса, а после активности – вибрациям Духовного Манаса, как уже отмечалось. Отсюда следует, что на этот отдел в равной степени может влиять, как гипофиз и средний мозг с мозжечком, так и эпифиз. Из цветов ему соответствуют градация зелёного и индиго, а из звуков – ноты Фа и Ля. В таламусе найдены 42 ядерных центра; выше приводится перечисление отделов головного мозга с ядрами и поэтому останавливаться на этом не имеет смысла. Ядра таламуса произвольно можно связать в определённые группы:
1. Передняя группа,
2. Латеральная группа,
3. Центральная группа,
4. Медиальная группа,
5. Вентральная группа,
6. Задняя группа.
В каждую из этих групп схематически можно включить по семь ядер, что даёт нам 42 ядра в шести ядерных центрах. Седьмым центром будет сам таламус, как собирательный и синтетический отдел. Данное схематическое распределение, несмотря на некоторую небрежность, позволит в дальнейшем более точно рассматривать, как звуковой, так и хроматический аспекты взаимодействий между таламическими и мозжечковыми центрами. Интересен тот факт, что между цветовыми, звуковыми вибрациями и иерархией ядер таламуса существует прямая связь. В связи с этим, следует привести отрывок из «Тайной Доктрины» Е.П.Блаватской. В частности, философ пишет: «При исследовании скорости вибраций цветовых вибраций обнаруживается, что каждый цвет отличается от последующего на шаг, равняющийся 42 (нм), или 6x7.
462 Красный + 42=504
504 Оранжевый + 42=546
546 Жёлтый + 42=588 Третья Октава восприятия психических цветов
588 Зелёный + 42=630
630 Синий + 42=672
672 Индиго + 42=714
714 Фиолетовый + 42=756
756 Красный
Проводя этот процесс в обратном порядке и вычитывая по 42, мы находим, что первым или основным цветом для нашего земного шара является зелёный цвет.
- Зелёный
42 Синий Первая полу октава
84 Индиго
126 Фиолетовый
168 Красный
210 Оранжевый Вторая октава
252 Жёлтый
294 Зелёный
336 Синий
378 Индиго
420 Фиолетовый
462 Красный».
Далее, великая Блаватская пишет о семеричном переходе в звуковой иерархии, начиная отсчёт со второй октавы, до первой полуоктавы; от ноты До – 539 второй октавы, до ноты Фа первой полуоктавы, как начальной на Земле. Ниже будет представлена полная схема гаммы переходов семи нот для одной октавы, от 49 до 343 переходов в соответствии с атомными подуровнями.
Утверждать прямо о принадлежности к той или иной хроматической и фоновой градациям тех или иных центров таламуса можно на данном этапе развития физиологии лишь интуитивно. То, что такая связь есть, это бесспорно, но определить данные связи, даже эмпирически и интуитивно очень сложно, хотя бы в силу того, что многие центры самого таламуса недостаточно изучены в аспекте функциональности. Поэтому здесь следует представить общие положения взаимосвязей в хроматическом и в звуковом аспектах между ядрами мозжечка и таламическими ядрами. Как уже отмечалось выше, мозжечок, являясь фактическим водителем ритма мозга, соответствует ноте До и красному цвету, тогда как таламус, относясь к межуточному мозгу, является носителем сдвоенного аспекта различных функций, как низших, так и высших. Между прочим, данное носительство является одним из основных в целом ряде и является базисом для сложной релейной программы самого таламуса. Через данный отдел мозга производится как бы центральное перепрограммирование: переключение от низлежащих отделов к отделам выслежащим и наоборот. При этом в таламусе протекают преобразовательные процессы, т.е. процессы, связанные с адаптацией проведения вибраций в различные отделы. Таким образом, если мозжечок, как утверждали справедливо Древние, является носителем всех сил и водителем первоначального, если, конечно, так можно выразиться, ритма, то таламус - координатор мозжечка и, тем самым, центральный координатор в головном мозге. Если подойти к изучению данного вопроса в аспекте философском и этическом, то взаимосвязь таламуса и мозжечка можно представить как взаимосвязь Мысли и Воли. Как без Воли не проявляется Мысль, так и без Мысли Воля была бы стихийным самоуничтожающим явлением. Здесь ко всему нужно добавить, что таламус посредством связи с эпифизом, гипоталамусом и гипофизом взаимосвязывает две системы - ЦНС и Эндокринную систему - в единый комплекс. Таким образом, таламус является в ЦНС как носителем вибраций Кама Манаса, так и носителем Духовного Манаса. Отсюда следует, что без данного отдела мозга не смогут существовать ни эпифиз, ни гипоталамус с гипофизом и наоборот. Таким образом, таламус может интегрировать сразу три хроматические и звуковые градации, а именно: Зелёную, Индиго и Жёлтую и, соответственно, ноту Фа, ноту Ля и ноту Ми[56], но об этом уже говорилось выше. Исходя из вышесказанного о таламусе, можно предположить, что мозжечок, соответствующий Каме (красный цвет и нота До), взаимосвязанный с ядрами таламуса, таким образом, дополняет деятельность последнего в нисходящем проведении, тогда как сам таламус дополняет интегрирующую роль мозжечка в аспекте восходящем. На нашей земной градации, в которой ныне пребывает большая часть человечества, господствующей нотой, как уже говорилось, является градация ноты Фа и соответствующий ей зелёный спектр призматических цветов. Таким образом, если рассматривать взаимосвязь между этими двумя отделами мозга в музыкальном и цветовом аспектах, нетрудно уявить нотную и хроматическую корреляции между нотами До и Фа, и между красным и зелёным цветами. Так как во взаимосвязях ядер мозжечка и таламуса, так же как и с другими отделами мозга наличествует реципрокность (возвратность связей в том или ином отделе мозга к тем отделам, от которых данный отдел связи принимает), то корреляция в звуковом аспекте может меняться то в сторону До, то в сторону Фа, плюс интервальные соотношения между ними.
Интервальные соотношения могущие быть между мозжечком и межуточным мозгом
1. 2 т. Б. 3 Do-Mi, Do-Fab-------------------------- Мозжечок – эпифиз и желудочки - мозжечок – средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте - эпифиз и желудочки.
2. 21/2 т. Ч. 4 Do-Fa, Do-Mi#----------------- Мозжечок – средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте, мозжечок – эпифиз и желудочки - средний мозги.
3. 3 т. Ув.4 Do-Fa#, Do-Solb---------------------- Мозжечок – средний мозг таламус и межуточный мозг без эпифиза в низшем аспекте - кора, мозжечок – кора - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте.
4. 3 т. Ум. 5 Do-Solb, Do-Fa#--------------------- Мозжечок – кора-средний мозг, мозжечок - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте - кора.
5. 31/2 т. Ч. 5 Do-Sol, Do-Labb-------------------- Мозжечок – кора, мозжечок – Межуточный - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте (La дважды понижается).
6. 4 т. М. 6 Do-Lab, Do-Sol#---------------------- Мозжечок – межуточный мозг в высшем аспекте -кора, мозжечок – кора - продолговатый мозг.
7. 41/2 т. Б. 6 Do-La, Do-Sibb--------------------------- Мозжечок – межуточный мозг, в высшем аспекте, мозжечок – лимбическая кора - кора (Si дважды понижена).
8. 5 т. М. 7 Do-Sib, Do-La#------------------------------------ Мозжечок – лимбическая система - межуточный мозг, мозжечок – межуточный мозг в высшем аспекте - лимбическая система.
К вышеуказанным интервальным соотношениям мы ещё вернёмся, теперь, продолжая тему, следует привести некоторые общие числовые коррелятивные соотношения между ядрами таламуса и центральными ядрами мозжечка. Как уже указывалось, в таламусе насчитывается 42 ядра, которые тесно взаимосвязаны с четырьмя ядрами мозжечка. Дабы найти математическое соотношение между ними необходимо привести отношение 42-х к 4-м: 42/4=10,5. Стало быть, на каждое мозжечковое ядро приходится 10,5 ядер таламуса, по соотношению. Таким образом, два таламических ядра будут связываться ядрами мозжечка. Схематически это выглядит так:
Рис. 16. Схематическое представление таламо-мозжечковых ядерных связей.
Объяснение в тексте.
Учитывая тот факт, что в головном мозге отделы парные (по закону симметрии), 84 ядра будут взаимодействовать с 8-ю ядрами мозжечка. Но это лишь внешняя часть анализа, которая, в сущности, ничего не поясняет, даже физиологу. Под данными внешними связями кроется один из самых сложных философско-математических анализов, восходящих к Древней Тайной Доктрине, т.н. Тетраграмматон или Тетраксис Пифагора. Поэтому вновь обратимся к вышеуказанным исчисленьям. Число 10,5 является половиной числа 21 (7 х 3). Данное число является составным числом древнего еврейского имени Иегова(х), в его написании на иврите, и разлагается в своём числовом значении на 10, 6, 5. В Священном Имени данная числовая комбинация удвоена, исходя из закона Аналогии Гермеса Трисмегиста и закона симметрии, удвоим эту комбинацию, что в комбинированной сумме нам даёт 20,12,10 или число 42. Это число полностью соответствует числу ядер таламуса. Число 10,5 в эзотерическом разночтении может быть изображено и как 105 (105 х 2=210), и как 501; простой арифметический закон гласящий, что от перестановки мест слагаемых значение суммы не меняется, применялся Древними в его скрытом значении. Таким образом, разделив число 42 на 4 мы получаем то число таламических ядер, которое связано с одним ядром мозжечка. Это число ключевое центральное и для числового значения таламических ядер, и для ядер мозжечка. В связи с этим нелишним будет упомянуть о расчётах, приводимых графиней Е.П.Блаватской во 2-м томе «Тайной Доктрины»:
«Это относится к процессу нисхождения в Огне на Вершину для создания человека и т.д. и объяснено лишь, как проверка и употребление чисел гор: ибо, с одной стороны, мы имеем 10+5+6=21 вниз к середине 501, и на другой стороне 6+5+10=21».
Таким образом, данное математическое положение чётко поясняет, как нисходящий связи таламуса с мозжечком, так и восходящие связи мозжечка к таламусу[57]. Отсюда следует вывод о том, что головной мозг есть точное подобие Космоса, где Единый Закон проявляется, как в галактике, так и в человеке.
После рассмотренного философско-математического анализа вновь вернёмся к музыкальной и хроматической динамикам во взаимоотношениях мозжечка и таламуса.
2 т. Б. 3 Do-Mi, Do-Fab-------------------------- Мозжечок – эпифиз и желудочки - мозжечок – средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте - эпифиз и желудочки.

21/2 т. Ч. 4 Do-Fa, Do-Mi#----------------- Мозжечок – средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте, мозжечок – эпифиз и желудочки - средний мозг.
3 т. Ув.4 Do-Fa#, Do-Solb---------------------- Мозжечок – средний мозг таламус и межуточный мозг без эпифиза в низшем аспекте - кора, мозжечок – кора - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте.

3 т. Ум. 5 Do-Solb, Do-Fa#--------------------- Мозжечок – кора - средний мозг, мозжечок - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте - кора.
![]()
31/2 т. Ч. 5 Do-Sol, Do-Labb-------------------- Мозжечок – кора, мозжечок – Межуточный - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте (La дважды понижается).
![]()
4 т. М. 6 Do-Lab, Do-Sol#---------------------- Мозжечок – межуточный мозг в высшем аспекте - кора, мозжечок – кора - межуточный мозг без эпифиза.
![]()
41/2 т. Б. 6 Do-La, Do-Sibb--------------------------- Мозжечок – межуточный мозг, в высшем аспекте, мозжечок – лимбическая кора - кора (Si дважды понижена)
![]()
5 т. М. 7 Do-Sib, Do-La#------------------------------------ Мозжечок – лимбическая система - межуточный мозг, мозжечок – межуточный мозг в высшем аспекте - лимбическая система.
Так выглядит хроматическая динамика в соответствии с музыкальными интервалами, при взаимоотношениях таламуса и ядер мозжечка, в целом:
![]()
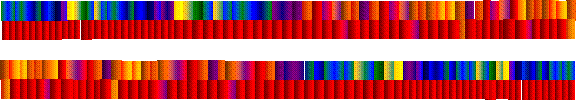
Как видно из данной спектральной диаграммы, при взаимодействии мозжечка с таламусом и всем межуточным мозгом, в хроматическом аспекте красному спектру мозжечка сопутствует широкий спектр от жёлтого до фиолетового, причём превалирующими цветами в данной шкале являются зелёный и сине-голубой. Первый представлен почти во всем своём спектре, от жёлто-зелёного до сине-зелёного. В этой хромодинамике заложен всё тот же семеричный принцип построения при одном Едином объединяющем. В музыкальном аспекте данным объединительным качеством обладают два тритона: увеличенная кварта и уменьшенная квинта, а в хроматическом аспекте – цветовая комбинация красного и сине-зелёного цветов. Эти 14 градаций, которые объединены двумя, представляют собой иерархию восходящих и нисходящих взаимосвязей мозжечка с межуточным мозгом и межуточного мозга с мозжечком, что в целом подтверждает вышеприведённый математический анализ из «Тайной Доктрины».
Замечу, что представленный ныне анализ, так же как и последующие, не является тонким и скрупулезным, но дан в общих философских аспектах. Тончайший органический и функциональный анализ требует написания целого тома и войдёт во вторую часть «Аналогий», «Частные аналогии».
Общая фоно-хроматическая картина взаимосвязи мозжечка со средним мозгом и с базальными ганглиями
Взаимосвязи среднего мозга с мозжечком, в аспекте музыки и хромодинамики, необходимо рассматривать в свете взаимоотношений Камы (Принципа Воли) и Кама Манаса (Животного Ума). Отсюда следует, что взаимоотношения будут между красной и зелёной цветовыми градациями, в хроматическом аспекте, и между градациями нот До и Фа, в музыкальном аспекте. В данных корреляциях, как и в корреляциях между таламусом, межуточным мозгом и мозжечком, принимают участие почти все отделы мозга. Вновь рассмотрим общие положения взаимосвязей мозжечка, но со средним мозгом.
2 т. Б. 3 Do-Mi, Do-Fab-------------------------- Мозжечок – эпифиз и желудочки - мозжечок – средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте- эпифиз и желудочки.
21/2 т. Ч. 4 Do-Fa, Do-Mi#----------------- Мозжечок – средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте, мозжечок – эпифиз и желудочки - средний мозг.
3 т. Ув.4 Do-Fa#, Do-Solb---------------------- Мозжечок – средний мозг таламус и межуточный мозг без эпифиза в низшем аспекте - кора, мозжечок – кора - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте.
3 т. Ум. 5 Do-Solb, Do-Fa#--------------------- Мозжечок – кора - средний мозг, мозжечок - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте - кора.

31/2 т. Ч. 5 Do-Sol, Do-Labb-------------------- Мозжечок – кора, мозжечок – Межуточный - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте (La дважды понижается).
![]()
В вышеуказанной спектральной диаграмме показан почти весь спектр зелёного цвета, ключевым уровнем в котором является чистый зелёный цвет, который объединяет три сопутствующих при данной корреляции цвета: жёлтый, голубой или чистый синий и индиго. Таким образом, вновь показана тесная взаимосвязь между двойственным Манасом и Буддхи. Данный анализ ещё один раз подтверждает тот факт, что мозжечок отдельно не интегрируется с тем или иным отделом, но вовлекает при этом все отделы мозга.
Общая фоно-хроматическая динамика во взаимосвязи мозжечка с лимбической системой
Лимбическая система, являющаяся одним из самых скрытых и малоизученных отделов в головном мозге, несёт в себе отражательные, экранные функции и, таким образом, способствует накоплению идеаций. Данную способность ряда отделов старых систем, например, гиппокампа, многие современные исследователи именуют глубинной памятью. К принципу памяти мы обратимся ниже, но данную способность лимбической структуры - накоплять и экранировать в те или иные отделы мозга различные идеации, наводит на мысль о соответствии этой системы с Астральным Двойником или Астральным телом в человеке, которое в древности именовали ещё зеркальным двойником.
41/2 т. Б. 6 Do-La, Do-Sibb--------------------------- Мозжечок – межуточный мозг, в высшем аспекте, мозжечок – лимбическая кора - кора (Si дважды понижена).
![]()
5 т. М. 7 Do-Sib, Do-La#------------------------------------ Мозжечок – лимбическая система - межуточный мозг, мозжечок – межуточный мозг в высшем аспекте - лимбическая система.
![]()
51/2 т. Б. 7 Do-Si, Do-Dob----------------------------------------- Мозжечок – лимбическая система, мозжечок – мозжечок-продолговатый мозг.
![]()
6 т. Ч. 8 Do-Do, Do-Si#, Do-Rebb--------------------------------- Мозжечок – мозжечок, Мозжечок – лимбическая система - мозжечок, мозжечок – продолговатый мозг - лимбическая система.
![]()
1/2 т. М. 2 Dob-Re, Do-Do# --------------- Мозжечок в нисходящей градации (мозжечок - лимбическая система) – продолговатый мозг, мозжечок – мозжечок - продолговатый мозг.
![]()
![]()
В данном случае нам представлен спектр от фиолетового индиго до оранжево-фиолетового и красно-оранжевого цветов, что говорит о наличие в данной корреляции как продолговатого мозга (оранжевый цвет, нота Ре), так и мозжечка (красный цвет, нота До). Здесь мы подходим к тесной взаимосвязи мозжечка с продолговатым мозгом и внутримозжечковым корреляциям. Данная корреляция, как и предыдущая, показывает как восходящие связи, так и нисходящие.
Общая фоно-хроматическая динамика во взаимосвязи мозжечка с продолговатым мозгом
Если мозжечок относится к Каме, а лимбическая система – к Астралу, то продолговатый мозг - где расположены многие жизненно важные центры: центр дыхания, рвотный центр, множество ядер и путей, связывающих головной и спинной мозги, ядра черепно-мозговых нервов и один из центров регуляции ритмом сердца, а также вегетативная нервная система и центры регуляции эндокринной системой, например, голубое пятно - должен соответствовать Пране – Жизненному Принципу на нашей планете.
51/2 т. Б. 7 Do-Si, Do-Dob----------------------------------------- Мозжечок – лимбическая система, мозжечок – мозжечок - продолговатый мозг.
![]()
6 т. Ч. 8 Do-Do, Do-Si#, Do-Rebb--------------------------------- Мозжечок – мозжечок, Мозжечок – лимбическая система- мозжечок, мозжечок – продолговатый мозг - лимбическая система.
![]()
0 т. Ч. 1 Do-Do, Dobb-Re-------------- Мозжечок – мозжечок, мозжечок дважды в нисходящей градации (мозжечок - эпифиз) – продолговатый мозг.
![]()
1/2 т. М. 2 Dob-Re, Do-Do# --------------- Мозжечок в нисходящей градации (мозжечок - лимбическая система) – продолговатый мозг, мозжечок – мозжечок - продолговатый мозг.
![]()
1 т. Б. 2 Do-Re, Do-Mibb------------------- Мозжечок – продолговатый мозг, Мозжечок – эпифиз и желудочки-мозжечок (нота Do дважды повышается).
![]()
11/2 т. М. 3 Do-Mib, Do#-Re---------------Мозжечок – эпифиз и желудочки - мозжечок продолговатый мозг – продолговатый мозг.
![]()
![]()
Общая прямая и опосредованная фоно-хроматическая динамика во взаимосвязи мозжечка с мозжечком
51/2 т. Б. 7 Do-Si, Do-Dob----------------------------------------- Мозжечок – лимбическая система, мозжечок – мозжечок - продолговатый мозг.
![]()
6 т. Ч. 8 Do-Do, Do-Si#, Do-Rebb--------------------------------- Мозжечок – мозжечок, Мозжечок – лимбическая система - мозжечок, мозжечок – продолговатый мозг - лимбическая система.
![]()
0 т. Ч. 1 Do-Do, Dobb-Re-------------- Мозжечок – мозжечок, мозжечок дважды в нисходящей градации (мозжечок - эпифиз) – продолговатый мозг.
![]()
1/2 т. М. 2 Dob-Re, Do-Do# --------------- Мозжечок в нисходящей градации (мозжечок - лимбическая система) – продолговатый мозг, мозжечок – мозжечок - продолговатый мозг.
![]()
1 т. Б. 2 Do-Re, Do-Mibb------------------- Мозжечок – продолговатый мозг, Мозжечок – эпифиз и желудочки - мозжечок (нота Do дважды повышается).
![]()
11/2 т. М. 3 Do-Mib, Do#-Re---------------Мозжечок – эпифиз и желудочки - мозжечок продолговатый мозг – продолговатый мозг.
![]()
![]()
Та же корреляция, что и выше, прослеживается и между взаимосвязями, как прямых, так и опосредованных, внутри мозжечка. Данной корреляции присущи, в основном, красно-оранжевые тона, которые свойственны Каме и Пране. Но корреляция показывает также и то, что внутримозжечковые связи не могут существовать отдельно, не вовлекая в этот единый корреляционный комплекс взаимосвязи с эпифизом и лимбической системой. Этот факт приводит к двум выводам:
1. Лимбическая система, соответствующая эфирному двойнику, в принципе, неразрывно связана с Камой – телом желаний.
2. Эпифиз, как носитель эманаций Духовной Души, или Буддхи, которая также связана с Праной, взаимосвязан и с продолговатым мозгом, по которому, в конечном итоге, поднимается к эпифизу Кундалини, и с мозжечком – носителем Воле-формы.
Общая прямая и опосредованная фоно-хроматическая динамика во взаимосвязи мозжечка с корой мозга
Кора головного мозга, как уже отмечалось, являясь конечным отделом, куда стекаются все идеации для синтеза и откуда уже в виде первичной информации, если так можно выразиться, направляются к низлежащим отделам мозга для завершения определённого цикла, соответствует Аурическому Яйцу, которое является синтезом всех эманаций тел в человеке.
3 т. Ув.4 Do-Fa#, Do-Solb---------------------- Мозжечок – средний мозг таламус и межуточный мозг без эпифиза в низшем аспекте - кора, мозжечок – кора - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте.
3 т. Ум. 5 Do-Solb, Do-Fa#--------------------- Мозжечок – кора - средний мозг, мозжечок - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте - кора.

31/2 т. Ч. 5 Do-Sol, Do-Labb-------------------- Мозжечок – кора, мозжечок – Межуточный - средний мозг, таламус и межуточный мозг без эпифиза в низшем аспекте (La дважды понижается).
![]()
4 т. М. 6 Do-Lab, Do-Sol#---------------------- Мозжечок – межуточный мозг в высшем аспекте - кора, мозжечок – кора - межуточный мозг без эпифиза.
![]()
41/2 т. Б. 6 Do-La, Do-Sibb--------------------------- Мозжечок – межуточный мозг, в высшем аспекте, мозжечок – лимбическая кора - кора (Si дважды понижена).
![]()
Мозжечково-корковая корреляция представлена холодным спектром цветов, основным цветом в котором является зелёно-синий или синий с индиго. Синий или голубой – цвет, который является цветом синтезированным из индиго и зелёного – цветов Манаса. В спектре данной корреляции отражен зелёный, т.к. это основной цвет мозга, как отражателя Низшего Манаса, он соседствует, как с голубым, так и с индиго – цветом Духовного Манаса. Фиолетовый цвет, соседствуя с синим (голубым), наводит на мысль о том, что в коре мозга, как и в лимбической коре отражаются все идеации от различных отделов мозга и всей ЦНС, в нашем земном физическом аспекте. Голубой же цвет проявляет себя при соединении Духовного Манаса с Буддхи – Духовной Душой.
Общий спектр цветовых корреляций между мозжечком и отделом головного мозга
Здесь, не представлены лишь корреляции мозжечка со спинным мозгом, т.к. последний,
являясь фактическим зеркальным отображением головного мозга в низшем аспекте, должен, по сути, не только выявлять взаимоотношения между собой и головным мозгом, но и вовлекать в данную корреляцию всю вегетативную и автономную нервные системы.
Примечателен и тот факт, что хроматическая корреляция, на показанном примере, аналогична т.н. «пачечной» активности, возникающей, например, при раздражении промежуточных нейронов (интернейронов), а также проявляющейся при фоновой импульсной активности нейронов. Относительно звуковых или т.н. шумовых возбуждений в ЦНС при «спонтанной» активности, которую не следует путать с фоновой активностью[58], следует отметить, что данные «шумовые», распространяющиеся возбуждения, есть ничто иное, как постоянная звуковая или музыкальная активность самих нейронов, которая всецело зависит как от структуры ЦНС, так и от глии. Как правильно поясняют нейрофизиологи, «спонтанная» активность подобна броуновскому движению, с разницей в масштабах. В отличие от классического представления данного движения, нужно отметить, что это далеко не хаотический процесс, а, наоборот – стремящийся к порядку. В ЦНС данное упорядочение производится очень быстро, и называется в нейрофизиологии процессом самоорганизации, которое происходит уже в эмбриональном периоде.
Основные мозжечковые музыкальные лады
Мозжечок, имея четыре центральных ядра в белом веществе, подчиняется, тем самым, основным элементарным законам Тетраксиса.
![]()
Тетрахорды в музыке. Как известно, тетрахордом называется совокупность и, что не маловажно, взаимное соответствие между собой звуков, располагающимися по секундам в одной чистой кварте. Диатонические тетрахорды бывают трёх видов:
Ионийский тетрахорд, с малой секундой вверху,
Дорийский – с малой секундой посередине, Фригийский – с малой секундой внизу.
Кроме данных видов тетрахордов, существуют лады, состоящие из двух различных тетрахордов, которые в сути своей дают семиступенчатый диатонический лад. Также и между вышеизложенными тетрахордами существуют сочетания, дающие, в сумме, семинотную диатонику.
Миксолидийский лад: ионийский и дорийский, Эолийский лад: дорийский и фригийский.
![]()
Лидийский лад Ув.4 м.2 ион.
Из вышеуказанного явствует, что тетрахорды в сочетаниях друг с другом интегрируют все ноты октавы, со всеми тональностями. Таким же образом ядра мозжечка интегрируются, посредством циклических взаимодействий, как друг с другом, так и с различными отделами мозга, объединяя все отделы мозга в единый комплекс. Получая афферентацию от тех или иных образований, то или иное ядро, взаимодействуя с другим ядром как своей стороны, так и другой, автоматически вовлекает в этот циклический процесс те образования, от которых получает афферентацию. Тем самым, ядра, «окрашивая», уже при выходе через клетки Пуркинье и через свои эфференты, те или иные образования (проводимые ими функции) в мозжечковые тона, сами «окрашиваются» через афферентацию в оттенки данных образований мозга. Данное положение подтверждает тот факт, что ни одно образование в мозге, как и во всей ЦНС, автономно не работает и не является автономным продуцентом той или иной функции. Нужно отметить также, что вибрации от афферентов в мозжечковые ядра, порождают целую гамму хроматизмов в работе мозжечка, при общей тонике До. Образованные при этом петли взаимодействия также могут способствовать выявлению тех же музыкальных интервалов и математических соотношений между мозжечком и остальным мозгом.
Несмотря на тот факт, что сам мозжечок, являясь тоническим отделом, не может быть единственным в подобном принципе, необходимо отметить, что в ЦНС все отделы имеют свои «дублирующие аналоги»; подобным «аналогом» для мозжечка является гиппокамп, который, как отмечалось выше, распределяет основной тон, поддерживая, тем самым, полное соответствие между всеми отделами и их образованиями. Как ни странно, но между самим гиппокампом и мозжечком не существует прямых (моносинаптических) связей в силу того, что оба отдела направлены на тоническое торможение. Так, если мозжечок тормозит (гасит) высокие вибрации от остальных образований, то гиппокамп, получая обильную афферентацию, постоянно поддерживает тон, заданный мозжечком. Таким образом, если две тормозящие системы имели бы прямое взаимодействие, происходил бы эффект анигилляции между этими отделами. Данные подсистемы бывают взаимосвязаны посредством релейных и компараторных подсистем, к которым относятся таламус, оливарные комплексы, ретикулярная формация, что, в свою очередь, позволяет подсистемам - сенсорной, вестибулярной и моторной - безошибочно проводить функции.
Стало быть, вся ЦНС подразделяется на семь принципиальных подсистем:
· Тонико-интегрирующая подсистема:
1. мозжечок,
2. септум
3. гиппокамп.
· Релейно-компараторная подсистема:
1. таламус, с несаецифическими ядрами,
2. оливарные комплексы,
3. ретикулярная формация.
· Сенсорная подсистема:
1. центры сенсорных органов,
2. ядра продоговатого мозга,
3. миндалины,
4. эпифиз.
· Проводниково-накопительная подсистема:
1. водопроводная система желудочков и каналов,
2. эпифиз,
3. гипоталамус,
4. гипофиз.
· Вестибулярная подсистема:
1. ядра вестибулярного аппарата,
2. кортиев орган и отолитовый аппарат,
3. верхне-оливарный комплекс,
4. кохлеарное ядро.
· Моторная подсистема:
1. средний мозг,
2. мост с ядрами,
3. продолговатый мозг,
4. спинной мозг.
· Зеркальная подсистема:
1. Кора мозга (на внешний план),
2. лимбическая кора (цингулярная извилина), энторинальная кора (на внутренний план).
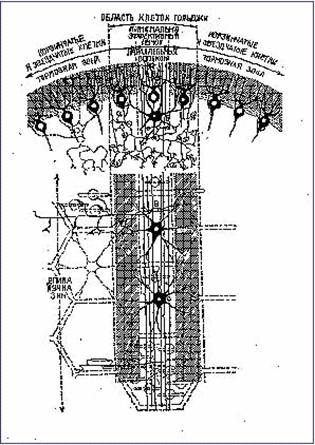
Рис. 17. Нейронная структура коры мозжечка с гексагональными схемами нейрональных сетей.
Сенсорная подсистема головного мозга на примере слухового и зрительного анализаторов и фоно-хроматические корреляции
Сенсорная подсистема анализа, являясь одной из разветвлёных в ЦНС, наряду с моторной и вестибулярной системами, взаимосвязывает организм с внешним миром. Любое изучение нервной системы, так или иначе связано с попытками определить степени данной взаимосвязи. Будет это электрофизиологическое исследование или исследование рефлекторных проявлений, исследования связанные с тонкими механизмами, протекающими в клетке и на её мембране, всё это лишь попытки определить максимальный спектр взаимоотношений живого организма с внешним миром. Одной из таких попыток является поиск взаимоотношений и взаимосоответствий между живым организмом и самыми яркими проявлениями Природы: звуком и цветом.
Ещё древние философы подметили, что всё, что ощущает человек, является лишь внешним проявлением истинного. Первые из известных нам философов, давшие определение чувствам и органам, воспринимающими эти чувства, были арии Северной Индии. В одном из древнейших научных трактатов Ариаварты – «Шатапатха-брахман» - есть достаточно точные представления о пяти чувствах человека. Следует отметить, что чувства, о которых идёт речь в вышеназванном трактате, рассматриваются не только в свете объективности и физического восприятия, но и в свете субъективного (внутреннего) восприятия данных чувств[59]. В дальнейшем античные философы, давая свои доктрины о строение человека, сделали попытку объединить физиологию и метафизику, основываясь на идее познания сущего посредством мысли и чувств.
Что кроется за чувствами? Откуда берётся источник самих восприятий? Не есть ли само восприятие одновременно и Узкими вратами в Вечность, и Широкими вратами в погибель? Эти вопросы много тысяч лет «стоят на повестке дня», а, между тем, проблема познания и самопознания остается актуальной. То, что мы видим, слышим, осязаем, обоняем и вкушаем, кроет под собой не только чисто физиологические механизмы в своих мельчайших подробностях, но и тончайшую взаимосвязь между органами восприятий и воспринимаемым в его подробностях. Именно посредством чувств мы по праву осознаём саму Вселенную, которая пребывает в нас и вне нас. Но для того, чтобы приблизиться к этому осознанию, необходимо уявить, что все природные законы и проявления ЕДИНЫ в своем МНОЖЕСТВЕ.
Слух и восприятия звука
«В Начале было Слово, и Слово было у Бога, и Слово было Бог». Так гласит первый стих Евангелия от Иоанна. Данное изречение не только сугубо религиозно, но и научно, ибо указывает на наличие первичной Идеи-Логоса – предтечи всего. Неслучайно, что именно данное выражение легло в начало нашего анализа слуха, т.к. звук как единица Материи является чуть ли не самой таинственной из всех форм той же Материи. Загадка, кроящаяся в природе звука, тесно связана с органами восприятия его, а также с головным мозгом, как системой анализирующей данное восприятие.
Строение слухового аппарата
Как известно, ухо млекопитающих состоит из наружного уха, среднего и внутреннего. Наружное ухо представляет собой ушную раковину и наружный слуховой проход, ведущий к барабанной перепонке. Далее следуют три мельчайших акустических косточки: молоточек, наковальня, стремечко, передающие колебания от барабанной перепонке к внутреннему уху. Последнее состоит их части вестибулярного лабиринта – улитки, содержащей в средней части базилярную мембрану, на которой расположены волосковые клетки. Процесс первичного поступления вибраций можно назвать этапом механической передачи акустических колебаний, где последним звеном, передающим данные волны является овальное окно и евстахиева труба. Как отмечает Г.Шеперд: «…чтобы звук стимулировал волосковые клетки, он должен быть сначала механически передан во внутреннее ухо, а затем надлежащим образом на волосковые клетки. Первый этап перехода звуковых волн из колебаний воздуха в колебание перелимфы. Это происходит посредством промежуточных движений косточек среднего уха. Поскольку воздух весьма сжимаем, а перелимфа несжимаема, косточки должны создать согласованность сил в этих двух средах; этот процесс называется согласованностью импендансов». Поглощая энергию с большой площади барабанной перепонки, косточки передают эту энергию на малую площадь стремечка, откуда она переходит (через овальное окно) на окружную мембрану улитки [Шеперд, 87]. Далее, как отмечает тот же автор, колебания передаются на базилярную мембрану улитки. Именно базилярная мембрана содержит волосковые клетки. Сама базилярная представляет собой пластинку, расширенную к узкой части улитки (апикальный конец) и суженную к широкой части улитки (базальный конец). Интересным представляется само устройство базилярной мембраны. Данная мембрана поперечно исчерчена суживающимися к базальному концу полосами, необходимыми, как это показал фон Гельмгольц для улавливания волн различной частоты. В дальнейшем Бекеши определил, что сами поперечные полосы по краям не расходятся, как если бы это были различной длины натянутые струны. Это подвело Бекеши к мысли о том, что при бегущей звуковой волне от овального окна к основанию мембраны, огибающая самой волны остаётся стационарной. Высокие по частотам пики смещаются к узкому концу мембраны, а низкие по частоте пики – к широкому концу, что было показано фон Гельмгольцем.
Как отмечалось, на базилярной мембране кортиева органа лабиринта улитки расположены волосковые клетки, подразделяющиеся на наружные и внутренние. Наружные, числом около 20000, своими верхними микроворсинками, так же как и внутренние, числом 3500, прикреплены к покровной мембране. Наружные волосковые клетки образуют три слоя, а внутренние – один. В 70-х годах прошлого столетья были проведены внутриклеточные опыты по определению акустических механизмов протекающих в самих клетках. Результатом явилось определение т.н. наилучшей или характеристической частоты настройки самих клеток. Было выявлено, что характеристической частотой для волосковых клеток является 17000 Гц. Те же данные, но гораздо раньше были получены Дэвисом и Галамбосом на отдельных волокнах слухового нерва. Нервные импульсы возникают синхронно с низкочастотными вибрациями до 1000 Гц. Звуковые частоты не кодируются только частотной импульсацией. Было определено, что сама частота кодируется положением клеток на базилярной мембране (тонотопическая организация).
Иннервация как наружных, так и внутренних волосковых клеток происходит следующим образом. Около 95% афферентных (чувствительных) волокон слухового нерва связаны с внутренними волосковыми клетками, тогда как 5% - с наружными. Эфферентные волокна, идущие от ядра верхней оливы, также образуют разветвлённые синаптические связи с внутренними и наружными волосковыми клетками. Причём полагают, что контралатеральных эфферентных связей больше, чем ипсилатеральных. Центробежные волокна способны угнетать ответоспособность волосковых клеток в целом [Шеперд, 1987]. Относительно малое число волокон слухового нерва (около 25000), соответствует такому же малому числу всех волосковых клеток. Об этой числовой взаимосвязи будет сказано ниже, в заключение здесь же следует вновь обратиться к работе Шеперда и указать на ряд тонких особенностей передачи звуковой импульсации к центральным отделам мозга.
Слуховые волокна, войдя в ствол мозга и дойдя до кохлеарного ядра, делятся на множественные ветвления. В кохлеарном ядре ветви эти строго упорядочены. Нужно отметить, что рецепторы базилярной мембраны также имеют широкое представительство в кохлеарном ядре.
Вторым пунктом в этой сложной специализации являются различные типы синапсов и клеток в ядре. Пфейфером была предложена интересная классификация клеток кохлеарного ядра, в основе которой лежит ответ на чистый тон различной частоты. Основной вывод кроется в том, что бифуркация первичных аксонов клеток ядра, а также специфичность синаптических связей, способны менять огибающую звуковой волны в самих первичных аксонах, на определённые ответы при выходе. Таким образом, каждый канал имеет свои клетки и свои места входов в отделы кохлеарного ядра. Также как и нейронная организация кохлеарного ядра, устроена организация взаимосвязей данного ядра со стволом мозга. Например, сферические клетки образуют двусторонние связи с верхнеоливарным комплексом; клетки-осьминоги, проецируются в тот же комплекс, но дают начало проекции оливы, через оливо-кохлеарный пучок, на улитку (центробежные волокна базилярной мембраны). Дорсальное кохлеарное ядро отдает проекцию в таламус, через нижнее четверохолмие. Уже на уровне таламических медиальных коленчатых тел, происходит релейная передача проекций на слуховую кору (первичную и вторичную) [там же].
Зрение и цвет
Зрение, являясь одним из ярких в своем проявлении чувств, столь же сложно и многогранно в строении, как и слух. Ориентированное на улавливание световых волн от всевозможных форм, органы зрения – глаза, явлены как самые выразительные части организма, способные выражать любое внутреннее проявление организма. Начиная с наружного органа зрения – глаза, нам открывается достаточно сложная, но строго упорядоченная система строения.
Как известно, глаз представляет собой сфероидную линзу, состоящую из белковой наружной оболочки, покрытого впереди роговицей, образующую радужную оболочку определённого цвета; ресничной связки; стекловидного тела, хрусталика (внутренняя двояковыпуклая линза); сетчатки с фоторецепторами, улавливающими фотоны; сосудистой оболочки; зрительного сосочка. Известно также, что проникновение световых лучей в глаз происходит по принципу рефракции. Свет, попадая в глаз, проникает на внутреннюю стенку глаза – сетчатку, на передней стороне которой расположены фоторецепторы: палочки и колбочки (у позвоночных). Кроме рецепторов на сетчатке расположены биполярные клетки, горизонтальные клетки, амакриновые и ганглиозные клетки. Первые (палочки) приспособлены улавливать основной темновой поток света, без распознавания цветов, тогда как вторые чувствительны к трём основным цветам спектра: красному, зелёному и синему. Вкратце, следует обратиться к самим механизмам фоторецепции. Фоторецепторы содержат мембранные диски, покрывающие реснички. Данные мембранные сегменты содержат фотопигмент родопсин, который и принимает одну из главных ролей в восприятии фотонов и переводе световой энергии в химическую. В этом сложном биохимическом процессе задействованы ионные каналы Na+, K+ и Ca2+, в качестве посредника. Так, Шеперд в той же монографии отмечает, что при темновом потоке и слабой освещённости, натриевые каналы бывают открыты, что приводит деполяризации мембраны. По одной из версий, как отмечает автор, это происходит благодаря cGMP-зависимому фосфорилированию белка Na+-канала. Изомерная форма родопсина, способствует ферментативному разложению cGMP, что приводит к обратному процессу – дефосфорилированию и закрытию натриевого канала. Не исключено, что изомерная форма родопсина запускает активность Ca2+, который приводит к закрытию натриевых каналов. По-видимому, эти два процесса происходят последовательно [Шеперд, 1987].
Относительно связи Na+-каналов со световосприятием следует отметить, что этот процесс может быть непосредственно связан с активностью эпифиза, т.к. известно, что именно эта железа начинает активизироваться ночью, а теряет активность именно в светлое время суток. Необходимо отметить также, что в регулировке Na+ и K+ активную роль также принимает шишковидная железа. Известно, например, что при недостаточности гипофиза (например, при надпочечниковой недостаточности), уровень натрия бывает повышен, что может указывать на «вынужденную» активацию со стороны эпифиза. Из вышеотмеченного, а также из-за того факта, что сам эпифиз является древним оптическим центром (Третий Глаз), видно, что ионное равновесие на сетчатке глаза может поддерживается благодаря шишковидной железе и ея взаимосвязи с гипоталамо-гипофизарной системой. Было бы неверным полагать, что один эпифиз осуществляет контроль за зрением. Чисто физиологически роль эпифиза в данном процессе столь же скрыта, сколь и общая его деятельность в мозге и в организме. С точки зрения классической физиологии, более яркую роль в процессе зрительного анализа отводят таламусу, верхним буграм четверохолмия, а также зрительной коре (затылочная область). Именно в эти отделы поступают зрительные пути, образующиеся от аксонов ганглиозных клеток сетчатки и сливающихся во вторую пару черепномозговых нервов – зрительных нервов. После зрительные нервы, формируя бинокулярный перекрёст, поступают в латеральные коленчатые тела таламуса. Проекции зрительного нерва до поступления к латеральным коленчатым телам поступают в верхние двухолмие, а также и в остальные отделы межуточного мозга. Однако исследования показали, что волокна нерва после двухолмия поступают в подушку таламуса, что позволяет судить о множестве параллельных путей от периферии к центру и о представительстве сетчатке в мозге (таламус) и в коре. Последним звеном в системе зрительного анализа является кора, а именно VI и VII (17-18) поля затылочной коры. Хьбелом и Визелом в 1974 году была предложена схема устройства по типу колонок глазодоминантности и ориентации зрительной коры. Функциональной единицей подобной колонки является определённый тип клеток: простые – воспринимающие лишь световые потоки, сложные – реагирующие на границу светового потока. Вышеотмеченные колонки имеют представительство в латеральных коленчатых телах [там же].
Выводы
Представленные выше данные являются классическим представлением об устройстве слуховой сенсорной подсистемы. Учитывая тот факт, что звук многомерен и, что самое главное, всепроникающ, напрашивается достаточно смелый вывод, относительно того, что абсолютно все отделы головного имеют акустические представительства в волосковых клетках. Было отмечено, что число волосковых клеток идентично числу волокон слухового нерва. 21000 наружных и 3500 внутренних волосковых клеток соответствуют 24500 волокнам слухового нерва. Разделив это число на 7 – число отделов мозга - мы получим 3500. Данное число – число волокон слухового нерва на каждый отдел мозга. Небезинтересно отметить, что наружные волосковые клетки, имеющие меньшее количество поступающих афферентных волокон от слухового нерва, играют очень важную роль в преобразовании звуковых волн в мозге. Они являются, своего рода преобразователями первичного звукового сигнала поступающего от внутренних волосковых клеток. Очень важно отметить, что данная фильтрующая система наружных волосковых клеток зависит, в первую очередь, от геометрического расположения последних на покровной мембране кортиева органа. Ниже представлена общая схема геометрического расположения наружных и внутренних волосковых клеток. Относительно данного трёхслойного расположения наружных волосковых клеток Шеперд отмечает, что богатая дивергенция волокон слухового нерва в этих клетках позволяет распространению поступающей импульсации от одной клетки к соседней, вовлекая тем самым в этот процесс все три ряда клеток последовательно. Тот же автор указывает на то, что основные звуковые ответы улавливаются внутренними волосковыми клетками, но, несмотря на эти утверждения, также говорится о том, что значение наружных волосковых клеток, равно как и механизмы ими осуществляемые, до конца не выяснены [там же]. Ответ на данную загадку кроется именно в дивергенции, о которой говорит Шеперд. Трёхслойное расположение наружных клеток, посредством вышеуказанной дивергенции, образует чёткие геометрические модули из 6-и клеток в каждом модуле с центральной клеткой. Данные модули с центром, являются взаимоперекрывающими, таким образом, что любая из клеток может являться центральной для своего модуля. Важно отметить и то предположение, что действующими на выход клетками являются именно 6 клеток, получающих импульсацию от центральной и передающих её соседним клеткам. Те клетки, которые для самих модулей являются центральными, не бывают вовлечены в выход данного, конкретного модуля. Следовательно, на выход из наружного слоя клеток всегда настроены именно 6 клеток, а не 7, как может показаться, при всём притом, что все клетки наружного слоя вовлечены в процесс получения звуковой импульсации клетками внутреннего слоя и передачи им импульсации по эфферентным путям от центральных отделов. Таким образом, огибающая звуковой волны в наружных волосковых клетках будет стационарна. Данная чёткость необходима для того, чтобы первичная звуковая импульсация была распределена от всех клеток внутреннего слоя, число которых 3500, наружным клеткам и наоборот. Следовательно, 21000 наружных волосковых клеток передают эфферентную импульсацию клеткам внутреннего слоя с максимальной точностью и отправляют уже преобразованную звуковую импульсацию спиральным ганглиозным клеткам второго порядка.
Вышеотмеченные выводы косвенно подтверждаются общим числом как волосковых клеток, так и волокон слухового нерва: 24500. Следует отметить, что волокна слухового нерва, имеющие чёткое константное числовое значение для определённого отдела мозга, не могут иметь подобного константного значения для различных образований в данных отделах, т.к. в каждом из отделов расположено различное количество образований. Следовательно, точное дискретное распределение волокон слухового нерва в головном мозге, должно соответствовать необходимости тех или иных образований в надёжной связи слуховой подсистемы с данными образованиями.

Рис. 18. Схема геометрического расположения наружных и внутренних волосковых клеток в кортиевом органе.
Как отмечалось выше в разделе о слухе и как поясняют сами физиологи, головной мозг получает от сенсорных органов материал для дальнейшей обработки путём прямых представительств периферии в центральных звеньях в иерархическом порядке. Как и в случае со слуховой системой, зрительная также имеет свои представительства в отделах мозга. Несмотря на общую схему представительств как для слуховой, так и для зрительной сенсорных систем в коре мозга, стволе и в таламусе, в схеме представительства системе рецептор-нервное волокно-центр, между слуховыми и зрительными периферийными центрами должны быть различия. Так, если рецепторные клетки кортиевого органа образуют вышеотмеченные модули по типу гексагона, то модули фоторецепторов устроены скорее по принципу клавиатуры рояля, т.к. цветовое восприятие всегда сопутствуется темновым потоком, который является основным. Модальной системой в сетчатке является группа следующих клеток: колбочки, палочки, биполярные клетки, горизонтальные клетки, амакриновые и ганглиозные клетки. Это нисколько не нарушает того факта, что зрительные рецепторные модули могут иметь свои представительства во всех отделах мозга, наподобие слуховых рецепторов. Общность в принципах представительского распределения в этих двух сенсорных системах, а также схожесть в устройствах самих систем позволяет говорить о том, что процессы восприятия как звука, так и света взаимозависимы, так же как и сами, звук и свет.
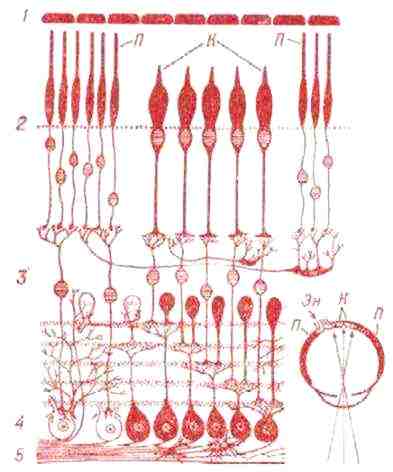
- · 1 - пигментные клетки - поглощают избыток света и транспортируют кислород, питающий сетчатку;
- · 2- светочувствительные клетки-фоторецепторы (П-палочки, обеспечивают сумеречное и ночное зрение;
- · К-колбочки, зрительное восприятие всей палитры цветов в дневное время, до 16 оттенков);
- · 3- двуполюсные нервные клетки;
- · 4- многополюсные нервные клетки;
- · 5- нервные волокна, отходящие от многополюсных клеток(из них состоит зрительный нерв).
Рис. 19. На схеме, справа, внизу, показано расположение палочек и колбочек на сетчатке и ход световых лучей. Зрительный нерв (Зн) указан короткой стрелкой.
продолжение следует
_________________________
[56] Речь тут идёт только о ЦНС, т.к. эпифиз, гипофиз и частично гипоталамус, относятся также и к эндокринной системе.
[57] См. раздел “Тетраксис Пифагора в человеческом мозге” данного труда.
[58] Спонтанная активность постоянно присутствует в ЦНС без каких-либо раздражений, тогда как фоновая активность, регистрируемая, как и спонтанная на мембране нейрона, способна изменятся от степени раздражения, несмотря на то, что является фоном для всех волновых процессов в нейроне.
[59] «Шатапатха-брахман», X, 3, 3; “Шатапатха-Брахман”, X, 5, 3.
Ваши комментарии к этой статье
№31 дата публикации: 01.09.2007
copyright © грани эпохи